1 引言
捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制.
事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用.
Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用.
在前人研究自扩散影响的基础上, Kerner[29 ] 首次提出了交叉扩散. 与考虑扩散在物种内发生的自扩散系统不同, 含交叉扩散的系统能够描述捕食者与猎物物种之间的扩散. Jalilian 等[30 ] 研究了具有群体防御行为的交叉扩散捕食者-猎物系统中的 Hopf 分岔和 Turing 不稳定性, 检查了分岔周期解的稳定性和 Hopf 分岔的方向. Tulumello 等[31 ] 研究了含交叉扩散的捕食者-猎物反应扩散系统的图灵斑图的形成过程, 通过非线性分析和数值模拟, 预测且验证了斑图模式以及波的形状和速度. Wang 等[32 ] 研究了具有交叉扩散、Allee 效应和 Holling IV 型功能响应函数的捕食者-猎物系统的斑图动力学, 给出了系统 Turing 不稳定、Hopf 不稳定和 Turing-Hopf 不稳定的理论分析. Han 等[33 ] 提出了一种具有捕食者趋向性的反应扩散捕食者-猎物系统, 揭示了斑图的形成是由捕食者趋向性和捕食者扩散的共同作用驱动的. 目前, 对反应扩散系统的研究多局限于单一相互作用和功能响应函数的情形. 因此, 本文基于改进型 Leslie-Gower 系统, 考虑 Beddington-DeAngelis 功能响应函数, 研究恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用机制影响下系统的时空动力学, 力图深入探明实际捕食者-猎物系统的动力学行为.
(1) 提出了一种改进型 Leslie-Gower 反应扩散捕食者-猎物系统, 该系统考虑了多种相互作用机制和 Beddington-DeAngelis 功能响应函数;
(2) 对无扩散系统进行 Hopf 分岔与稳定性分析, 并以此为前提得到系统发生 Hopf 分岔与产生 Turing 不稳定的条件;
(3) 对扩散系统进行数值仿真, 阐明了不同相互作用强度和交叉扩散系数的改变对斑图模式的影响, 并且给出了不同斑图模式生物学意义上的解释; 且不同的相互作用机制还会在不同程度上改变系统稳定性和图灵斑图模式的稳定速度.
2 系统描述
根据文献 [34 ] 中的研究, 综合考虑改进的 Leslie-Gower 项和 Beddington-DeAngelis 功能响应函数, 能有效描述捕食者在不同种群密度下的捕食行为. 但并没有考虑捕食者-猎物之间广泛存在的相互作用和物种迁徙模式. 为了提高系统的普遍性和真实性, 在文献 [34 ] 的基础上, 加入恐惧效应、种内竞争等相互作用, 并进一步引入交叉扩散. 改进的系统模型如下
(2.1) $\begin{equation} \begin{cases}\dfrac{\partial u}{\partial t}=D_{u}\nabla^{2}u+D_{uv}\nabla^{2}v+\dfrac{ru}{1+fv}-du-hu^{2}-\dfrac{quv}{1+bu+cv},\\\dfrac{\partial v}{\partial t}=D_{vu}\nabla^{2}u+D_{v}\nabla^{2}v+v(\beta-\dfrac{mv}{u+\alpha}),&\end{cases} \end{equation}$
系统 (2.1) 遵循非负性条件 $ u(x, y, 0) \geq 0, \, v(x, y, 0) \geq 0, \, (x, y) \in \Omega $
其中, $\nabla^{2}$ $\Omega\subset R^2$ $\partial\Omega$ $ \boldsymbol{n} $ $\partial\Omega$ $u$ $v$ $r$ $d$ $h$ $\beta$ $m$ $f$ $q$ $b$ $c$ $\alpha$ $D_{u}>0$ $D_{v}>0$ $D_{uv}$ $D_{vu}$
$\textbf{注 2.1}\quad$ $D_{u}D_{v}-D_{uv}D_{vu}>0$ .
3 无扩散系统分析
3.1 共存平衡点
当系统 (2.1) 不存在扩散现象时, 退化的无扩散系统如下
(3.1) $\begin{equation} \begin{cases}\dfrac{{\rm d}u}{{\rm d}t}=\dfrac{ru}{1+fv}-du-hu^2-\dfrac{quv}{1+bu+cv}, & \$3mm] \dfrac{{\rm d}v}{{\rm d}t}=v\bigg(\beta-\dfrac{mv}{u+\alpha}\bigg). & \end{cases} \end{equation}$
通过计算得到系统 (3.1) 的共存平衡点为 $E^{*}(u^{*},v^{*})$ $v^*=\frac{\beta(u^*+\alpha)}{m}$ . 将 $v^*=\frac{\beta(u^*+\alpha)}{m}$
$A=-cfh\beta^{2}-bfhm\beta$
$B=-(fq+cdf+2cfh\alpha)\beta^{2}-(chm+fhm+bdfm+bfhm\alpha)\beta-bhm^{2}$
$\textbf{假设 3.1}\quad D>0$ .
$\textbf{定理 3.1}\quad$ $E^{*}(u^{*},v^{*})$ .
$\textbf{证}\quad$ $A<0,B<0$ . 若假设 3.1 成立, 则 $D>0$ $f(u)=Au^3+Bu^2+Cu+D$ $f\left(0\right)=D>0, \lim_{u\to+\infty}f(u)<0$ . 根据零点定理, 方程 (3.2) 存在正根. $f^\prime(u)=3Au^2+2Bu+C$ $f^{\prime}(u)=0$ $f^\prime(u)=0$ $u_{1,2}=\frac{-B\pm\sqrt{B^{2}-3AC}}{3A},$
(1) 当 $B^2-3AC\leq0$ $f^\prime(u)=0$ $ f^{\prime}(u)$ $f(u)$
(2) 当 $B^2-3AC>0$ $B^2-3AC>B^2$ $f^{\prime}(u)=0$ $u_+$ $u_-$ $f(u)$ $(0,u_{+})$ $(u_{+},+\infty)$
(3) 当 $B^2-3AC>0$ $B^2-3AC<B^2$ $f^{\prime}(u)=0$ $f(u)$ $(0,+\infty)$
因此系统 (3.1) 在给定的系统参数均为正常数且满足假设 3.1 的条件下, 存在唯一正平衡点 $E^{*}(u^{*},v^{*})$ .
3.2 稳定性分析
下面计算共存平衡点 $E^{*}(u^{*},v^{*})$
(3.3) $\lambda_{0}^{2}-M_{1}\lambda_{0}+M_{2}=0,$
$\textbf{定理 3.2}\quad $ $E^{*}(u^{*},v^{*})$
$\textbf{证}\quad$ $M_1<0$ $M_2>0$ $E^*(u^*,v^*)$
$\textbf{注 3.1}\quad$
3.3 Hopf 分岔分析
选参数 $h$ $h_H.$ $\lambda_0=\eta+{\rm i}\phi$ $h=h_{H}$ $\lambda_0={\rm i}\phi$
$\textbf{定理 3.2}\quad$ $h=h_{H}$
$\textbf{证}\quad$ $g(\lambda_{0})=\lambda_{0}^{2}-M_{1}\lambda_{0}+M_{2}$ $h$
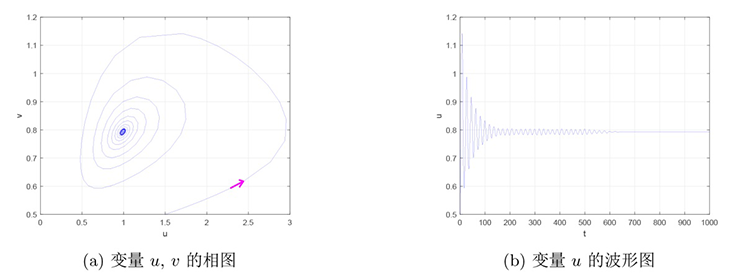
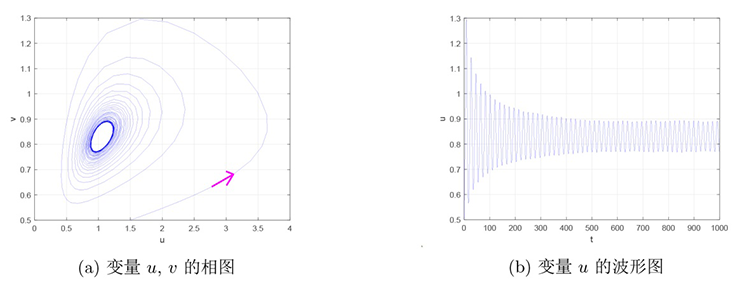
故而当假设 3.3 成立时,当参数 $h$ $h_{H}$ $[\lambda_0]$ $r=1.4,d=0.1,f=0.8,q=1.8,b=1.2,c=0.2,\alpha=0.2,\beta=0.2,m=0.3$ $h$ 图1 和图2 所示的相位图和波形图. 图1 表明当 $h=0.15>h_{H}$ $E^{*}$ 图2 表明当 $h=0.12<h_{H}$ $E^{*}$
图1
图1
$h{=}0.15{>}h_{H}$ $E^{*}$
图2
图2
$h{=}0.12{<}h_{H}$ $E^{*}$
4 空间扩散系统分析
4.1 Turing 不稳定
(4.1) $\begin{pmatrix}u\\v\end{pmatrix}=\begin{pmatrix}u^*\\v^*\end{pmatrix}+\begin{pmatrix}\epsilon_1\\\epsilon_2\end{pmatrix}\exp\left(\lambda_kt+{\rm i}k\cdot\boldsymbol{\tau}\right),$
其中, $ 0<\epsilon_{1},\epsilon_{2}\ll1,\lambda_{k}$ $t$ $\boldsymbol{\tau}$ $k>0$ $k^{2}=k_x^{2}+k_y^{2},k\cdot\boldsymbol{\tau}=\left(k_{x}x+k_{y}y\right).$
在共存平衡点 $E^{*}$
(4.2) $\begin{cases}\dfrac{\partial u}{\partial t}=J_{11}u+J_{12}v+D_{u}\nabla^2u+D_{uv}\nabla^2v, &\\[3mm]\dfrac{\partial v}{\partial t}=J_{21}u+J_{22}v+D_{vu}\nabla^2u+D_{v}\nabla^2v. & \end{cases}$
将方程 (4.1) 代入系统 (4.2), 可以得到系统 (4.2) 在平衡点 $E^{*}$
从而得到关于 $\lambda_{k}$
(4.3) $\lambda_k{}^2+G\left(k^2\right)\lambda_k+H\left(k^2\right)=0,$
$\textbf{定理 4.1}\quad$ $E^{*}$
$\textbf{证}\quad$ $G(k^2)>0$ $k$ $H(k^2)<0$ . 根据 Routh-Hurwitz 判据,此时特征方程 (4.3) 存在正实部特征根, 即系统 (2.1) 在平衡点 $E^{*}$ $H(k^2)$ $k^2$ $k_\mathrm{min}$
(4.4) $k_{\min}=\left(\frac{D_{v}J_{11}+D_{u}J_{22}-D_{uv}J_{21}-D_{vu}J_{12}}{2\det(D)}\right)^{\frac{1}{2}},$
将 (4.4) 式代回到 $H(k^2)$ $H(k^2)<0$
因此如果假设 3.2 和假设 4.1 成立, 那么系统 (2.1) 在平衡点 $E^{*}$
5 数值模拟
5.1 不同相互作用类别和强度改变系统空间格局
本小节是在二维平面上对系统 (2.1) 的空间斑图进行数值模拟. 在 $n\times n=140\times140$ $dx=dy=0.5$ $dt=0.01$ $t=1000.$
其中, rand$(n)$ $n\times n=140\times140$ $0\sim1$ $r=1.4,d=0.1,c=0.2,q=1.8,\alpha=0.2,\beta=0.2,D_{u}=0.02,D_{v}=2,D_{uv}=0.05,D_{vu}=0.005.$
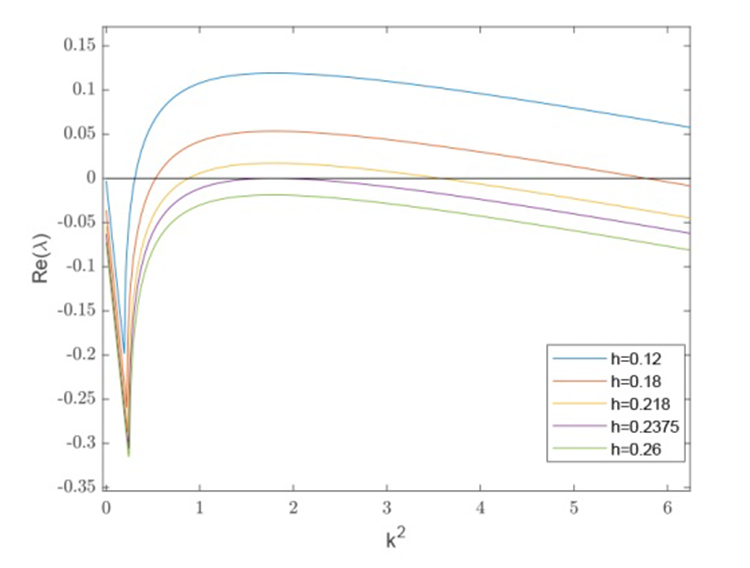
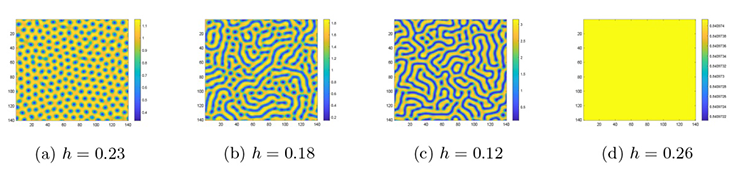
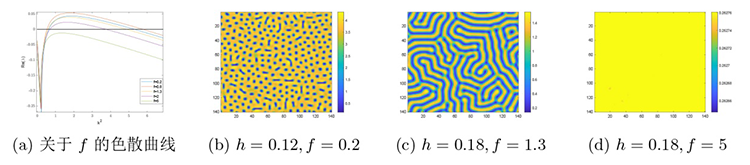
首先, 设定其余参数为 $f=0.8,b=1.2,m=0.3$ . 通过变化 $h$ 图3 所示的色散曲线及图4(a)-(d) 所示的不同斑图模式. 当 $h=0.23$ 图4(a) ), 猎物种群形成稳定且规则的点状斑图. 当 $h$ $h=0.18$ 图4(b) ), 点与条状斑图的分布整体趋于平衡, 当 $h=0.12$ 图4(c) ), 斑图模式转变为以条状斑图占主导的迷宫斑图, 而当 $h=0.26$ 图4(d) ), 斑图模式转变为纯色斑图, 即系统稳定性发生了变化.
图3
图4
图4
系统 (2.1) 在猎物种内竞争系数 $h$
$\textbf{注 5.1}\quad$ 图4 斑图模式变化的解释: 当猎物种内竞争系数 $h$ $h$ $h$
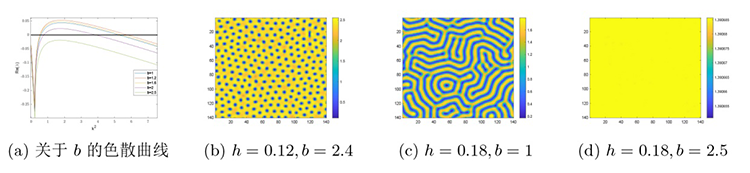
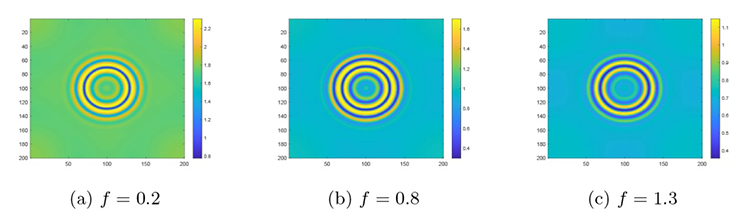
接下来是在对猎物种内竞争研究的基础上讨论恐惧效应对斑图模式及系统稳定性的影响. 参数保持不变, 通过变化 $h$ $f$ 图5(a) 所示的色散曲线与图5(b)-(d) 所示的斑图模式. 图5(a) 是设定 $h=0.18$ $f$ 图5(b) 是设定 $h=0.12$ $f$ 图5(c) 设定 $h=0.18$ $f$ 图5(d) 设定 $h=0.18$ $f$ $h$ $f$
图5
图5
系统 (2.1) 在恐惧效应系数 $f$
$\textbf{注 5.2}\quad$ 图4 到图5 斑图模式转变的解释: 当恐惧效应系数 $f$ $f$
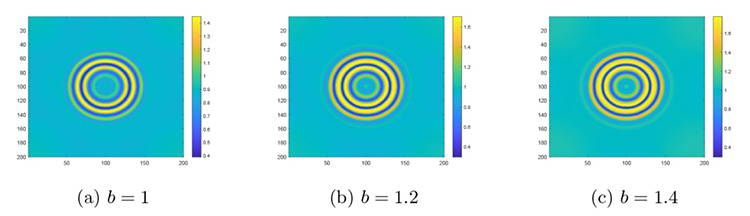
然后是在对猎物种内竞争研究的基础上讨论捕食者捕食猎物的饱和效应对斑图模式及系统稳定性的影响. 参数保持不变,通过变化 $h$ $b$ 图6(a) 所示的色散曲线与图6(b)-(d) 所示的斑图模式. 图6(a) 是设定 $h=0.18$ $b$ 图6(b) 是设定 $h=0.12$ $b$ 图6(c) 设定 $h=0.18$ $b$ 图6(d) 设定 $h=0.18$ $b$ $f$ $h$ $b$ $f$ $h$
图6
图6
系统 (2.1) 在饱和效应系数 $b$
$\textbf{注 5.3}\quad$ 图4 到图6 斑图模式转变的解释: 根据文献 [35 ,36 ] 的研究, 捕食者对猎物的饱和效应本质上反映了捕食过程中的时间成本约束. 捕食者从捕获猎物到完成进食、消化都需消耗一定时间资源, 这导致其单位时间内可处理的猎物数量存在上限. 当 $b$ $b$
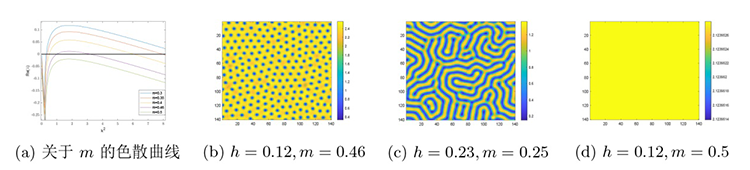
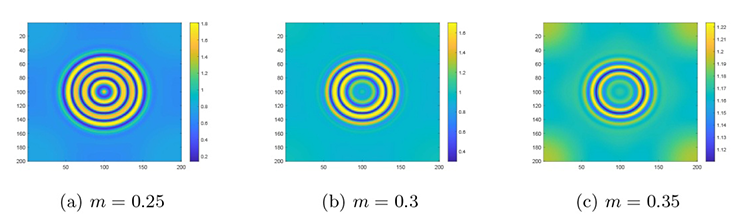
最后是在对猎物种内竞争研究的基础上讨论捕食者种内竞争对斑图模式及系统稳定性的影响. 参数保持不变, 通过变化 $h$ $m$ 图7(a) 所示的色散曲线与图7(b)-(d) 所示的斑图模式. 图7(a) 是设定 $h=0. 12$ $m$ 图7(b) 是设定 $h=0. 12$ $m$ 图7(c) 设定 $h=0. 23$ $m$ 图7(d) 设定 $h=0. 18$ $m$ $h$ $m$
图7
图7
系统 (2.1) 在捕食者种内竞争系数 $m$
$\textbf{注 5.4}\quad$ 图4 到图7 斑图模式转变的解释:当增大捕食者种内竞争系数, 捕食者个体之间的竞争压力加剧, 为减少因过度竞争而导致资源获取降低量, 每个捕食者在获取资源时倾向于避开其他捕食者以减少个体之间的竞争. 这种行为导致捕食者的分布趋于分散, 从而在捕食者分布的空隙区域, 猎物得以形成局部高密度聚集, 以降低被捕食的风险. 因此, 增大捕食者种内竞争系数会将条状斑图演化成点状斑图. 当减小捕食者种内竞争系数, 捕食者个体之间的竞争减弱, 捕食者个体更加广泛地分布到空间中, 这导致捕食压力在空间总体上变得更为一致, 从而导致猎物的空间分布更趋于均匀. 因此, 减小捕食者种内竞争系数会将斑图从点状斑图转变为条状斑图.
5.2 不同相互作用类别和强度改变斑图稳定速度
本小节首先通过考虑时空间离散域中猎物 $u$ $r=1.4, d=0.1, c=0.2, q=1.8, \alpha=0.2, \beta=0.2, D_{u}=0.02, D_{v}=2, D_{uv}=0.05, D_{vu}=0.005, f=0.8, b=1.2, m=0.3, h=0.18.$ 图8(a)-(d) 所示的关于四种不同相互作用及其强度对斑图演化到稳定的速度影响曲线.
图8
图8
系统 (2.1) 在不同相互作用及其强度下的斑图稳定速度曲线.
观察图8(a)-(d) 可知, 当稳定到平衡点浓度的网格个数不变, $h$ $f$ $m$ $h$ $m$ $f$ $b$ $n\times n=200\times200$ $t=100$
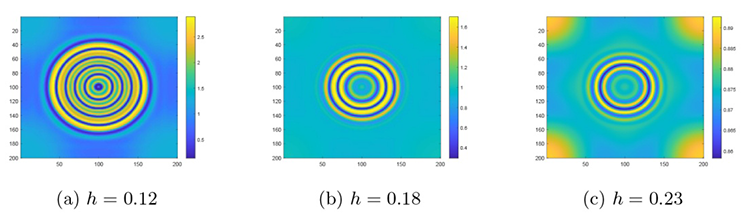
其余设置不变, 分别改变 5.1 小节讨论过的四种相互作用系数, 得到了如图9 -12 所示的关于四种不同相互作用及其强度下的斑图演化过程. 观察到斑图呈现一种环形波纹结构, 在此将其称作为环波, 通过相同时间下环波的数量不同来说明斑图的稳定速度.
图9
图9
系统 (2.1) 在不同 $h$
图10
图10
系统 (2.1) 在不同 $f$
图11
图11
系统 (2.1) 在不同 $m$
图12
图12
系统 (2.1) 在不同 $b$
观察图9 -12 可知, $h$ $f$ $m$ $h$ $m$ $f$ $b$
5.3 不同交叉扩散系数改变系统空间格局
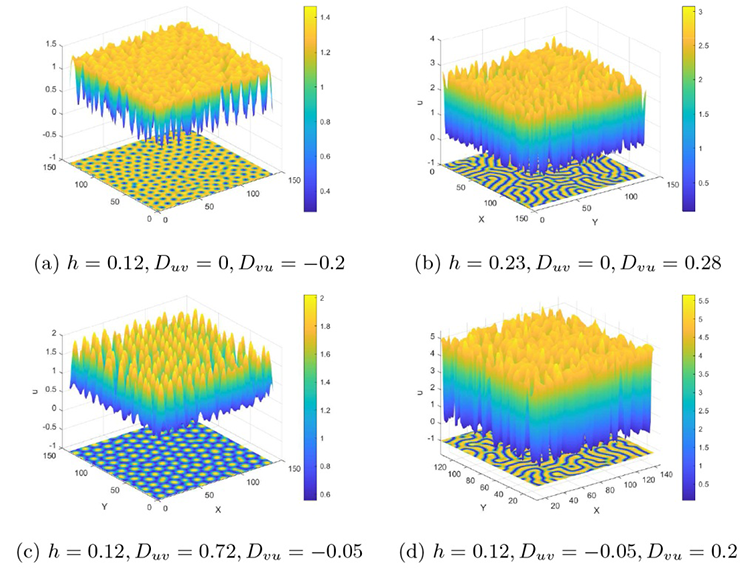
本小节在对猎物种内竞争研究的基础上讨论交叉扩散系数对斑图模式的影响. 其数值仿真的时空间初始条件设置与 5.1 小节一致, 参数设置为 $r=1.4,d=0.1,c=0.2,q=1.8,\alpha=0.2,\beta=0.2,D_{u}=0.02,D_{v}=2, f=0.8,b=1.2,m=0.3$ $h$ $D_{uv}$ $D_{vu}$ 图13(a)-(d) 所示的不同斑图模式. 这里引入三维视角, 以便于观察斑图的形成机理.
图13
图13
系统 (2.1) 在不同交叉扩散系数组合取值下的斑图.
可以看到, 不同的交叉扩散系数组合下, 系统 (2.1) 展现出不同的斑图模式. 仅通过改变交叉扩散系数, 可以让斑图模式从条状转变为点状, 还可以从点状转变为条状. 此外, 还生成了如图13(c) 所示的星空点状斑图. 其丰富的斑图模式表明, 交叉扩散对系统 (2.1) 具有重要影响. 因此, 在捕食者-猎物系统中引入交叉扩散是至关重要且不可忽视的.
6 结论
本文针对捕食者-猎物系统中广泛存在的多种相互作用及交叉扩散, 提出了一个较为综合的反应扩散捕食者-猎物系统, 该系统考虑了多种相互作用机制及交叉扩散因素, 研究了其无扩散系统的线性稳定性及其 Hopf 分岔, 得到无扩散系统在平衡点处的局部渐进稳定和发生 Hopf 分岔的条件. 对扩散系统研究了其发生 Turing 不稳定的条件. 通过绘制色散关系曲线, 结合数值模拟, 验证了不同相互作用机制和交叉扩散对捕食者-猎物动态行为的影响. 其数值结果表明, 种内竞争、恐惧效应、饱和效应以及交叉扩散都会引起斑图模式在点状、点条共存、条状斑图之间的相互转变. 不同的相互作用机制还会在不同程度上引起系统稳定性的转变并影响其斑图稳定速度. 本文的研究展示了广泛的时空动态特性, 这对研究捕食者-猎物的动态行为具有重要的理论和应用意义. 此外, 未来将开展以下工作来深化对捕食者-猎物动态行为的理解: 1) 引入三维位置信息, 研究系统 (2.1) 的三维空间斑图模式, 以捕捉更复杂的时空动态; 2) 加入控制策略, 研究控制器对系统 (2.1) 动态行为的作用, 探索如何通过外部干预实现对系统 (2.1) 的控制; 3) 进行振幅方程推导, 为斑图模式的形成与转变提供数学理论支撑.
参考文献
View Option
[1]
Pal D Kesh D Mukherjee D Cross-diffusion mediated Spatiotemporal patterns in a predator-prey system with hunting cooperation and fear effect
Mathematics and Computers in Simulation , 2024 , 220 128 -147
DOI:10.1016/j.matcom.2024.01.003
URL
[本文引用: 1]
[2]
Volterra V Variations and fluctuations of the number of individuals in animal species living together
ICES Journal of Marine Science , 1928 , 3 1 ): 3 -51
DOI:10.1093/icesjms/3.1.3
URL
[本文引用: 1]
[3]
Berryman A A The orgins and evolution of predator-prey theory
Ecology , 1992 , 73 5 ): 1530 -1535
DOI:10.2307/1940005
URL
[本文引用: 1]
Predator—prey theory is traced from its origins in the Malthus—Verhulst logistic equations, through the Lotka—Volterra equations, logistic modifications to both prey and predator equations, incorporation of the Michaelis—Menten—Holling functional response into the predator and prey equations, and the recent development of ratio—dependent functional responses and per—capita rate of change functions. Some of the problems of classical predator–prey theory, including the paradoxes of enrichment and biological control, seem to have been caused by the application of the principle of mass action to predator–prey interactions. Predator–prey models that evolved from logistic theory or that incorporate ratio—dependent functional responses do not have these problems and also seem to be more biologically plausible.
[4]
Gause G F Smaragdova N P Witt A A Further studies of interaction between predators and prey
The Journal of Animal Ecology , 1936 , 1 1 -8
[5]
Crombie A C On competition between different species of graminivorous insects
Proceedings of the Royal Society of London. Series B-Biological Sciences , 1945 , 132 869 ): 362 -395
[6]
Canale R P An analysis of models describing predator-prey interaction
Biotechnology and Bioengineering , 1970 , 12 3 ): 353 -378
DOI:10.1002/bit.v12:3
URL
[7]
Goel N S Maitra S C Montroll E W On the Volterra and other nonlinear models of interacting populations
Reviews of modern physics , 1971 , 43 2 ): 231 -276
DOI:10.1103/RevModPhys.43.231
URL
[本文引用: 1]
[8]
Seo G DeAngelis D L A predator-prey model with a Holling type I functional response including a predator mutual interference
Journal of Nonlinear Science . 2011 , 21 811 -833
DOI:10.1007/s00332-011-9101-6
URL
[本文引用: 1]
[9]
Antwi-Fordjour K Parshad R D Beauregard , M A Dynamics of a predator-prey model with generalized Holling type functional response and mutual interference
Mathematical biosciences , 2020 , 326
[本文引用: 1]
[10]
Jiang X She Z Ruan S Global dynamics of a predator-prey system with density-dependent mortality and ratio-dependent functional response
Discrete and Continuous Dynamical Systems , 2021 , 26 4 ): 1967 -1990
[本文引用: 1]
[11]
Cantrell R S Cosner C On the dynamics of predator-prey models with the Beddington-DeAngelis functional response
Journal of Mathematical Analysis and Applications , 2001 , 257 1 ): 206 -222
DOI:10.1006/jmaa.2000.7343
URL
[本文引用: 1]
[13]
王灵芝 . 具有恐惧效应的时滞捕食者-食饵模型
吉林大学学报 (理学版) , 2023 , 61 3 ): 449 -458
[本文引用: 1]
Wang L Z A predator-prey model with fear effect and time delay
Journal of Jilin University (Science Edition) , 2023 , 61 3 ): 449 -458
[本文引用: 1]
[14]
Pal S Majhi S Mandal S Pal N Role of fear in a predator-prey model with Beddington-DeAngelis functional response
Zeitschrift für Naturforschung A , 2019 , 74 7 ): 581 -595
DOI:10.1515/zna-2018-0449
URL
In the present article, we investigate the impact of fear effect in a predator–prey model, where predator–prey interaction follows Beddington–DeAngelis functional response. We consider that due to fear of predator the birth rate of prey population reduces. Mathematical properties, such as persistence, equilibria analysis, local and global stability analysis, and bifurcation analysis, have been investigated. We observe that an increase in the cost of fear destabilizes the system and produces periodic solutions via supercritical Hopf bifurcation. However, with further increase in the strength of fear, system undergoes another Hopf bifurcation and becomes stable. The stability of the Hopf-bifurcating periodic solutions is obtained by computing the first Lyapunov coefficient. Our results suggest that fear of predation risk can have both stabilizing and destabilizing effects.
[15]
Sarkar K Khajanchi S Impact of fear effect on the growth of prey in a predator-prey interaction model
Ecological complexity . 2020 , 42
[本文引用: 1]
[16]
Adler P B Smull D Beard K H et al . Competition and coexistence in plant communities: Intraspecific competition is stronger than interspecific competition
Ecology Letters , 2018 , 21 9 ): 1319 -1329
DOI:10.1111/ele.13098
PMID:29938882
[本文引用: 1]
Theory predicts that intraspecific competition should be stronger than interspecific competition for any pair of stably coexisting species, yet previous literature reviews found little support for this pattern. We screened over 5400 publications and identified 39 studies that quantified phenomenological intraspecific and interspecific interactions in terrestrial plant communities. Of the 67% of species pairs in which both intra- and interspecific effects were negative (competitive), intraspecific competition was, on average, four to five-fold stronger than interspecific competition. Of the remaining pairs, 93% featured intraspecific competition and interspecific facilitation, a situation that stabilises coexistence. The difference between intra- and interspecific effects tended to be larger in observational than experimental data sets, in field than greenhouse studies, and in studies that quantified population growth over the full life cycle rather than single fitness components. Our results imply that processes promoting stable coexistence at local scales are common and consequential across terrestrial plant communities.© 2018 John Wiley & Sons Ltd/CNRS.
[17]
Letten A D Stouffer D B The mechanistic basis for higher-order interactions and non-additivity in competitive communities
Ecology letters , 2019 , 22 3 ): 423 -436
DOI:10.1111/ele.13211
PMID:30675983
Motivated by both analytical tractability and empirical practicality, community ecologists have long treated the species pair as the fundamental unit of study. This notwithstanding, the challenge of understanding more complex systems has repeatedly generated interest in the role of so-called higher-order interactions (HOIs) imposed by species beyond the focal pair. Here we argue that HOIs - defined as non-additive effects of density on per capita growth - are best interpreted as emergent properties of phenomenological models (e.g. Lotka-Volterra competition) rather than as distinct 'ecological processes' in their own right. Using simulations of consumer-resource models, we explore the mechanisms and system properties that give rise to HOIs in observational data. We demonstrate that HOIs emerge under all but the most restrictive of assumptions, and that incorporating non-additivity into phenomenological models improves the quantitative and qualitative accuracy of model predictions. Notably, we also observe that HOIs derive primarily from mechanisms and system properties that apply equally to single-species or pairwise systems as they do to more diverse communities. Consequently, there exists a strong mandate for further recognition of non-additive effects in both theoretical and empirical research.© 2019 John Wiley & Sons Ltd/CNRS.
[18]
苏茹燕 , 杨文生 . 具有集群效应和种内竞争的双时滞捕食模型的稳定性与 Hopf 分支
应用数学 , 2022 , 35 3 ): 662 -679
[本文引用: 1]
Su R Y Yang W S Stability and hopf bifurcation of a dual-delay predator-prey model with grouping effect and intra-specific competition
Applied Mathematics , 2022 , 35 3 ): 662 -679
[本文引用: 1]
[19]
Souna F Lakmeche A Djilali S The effect of the defensive strategy taken by the prey on predator-prey interaction
Journal of Applied Mathematics and Computing , 2020 , 64 665 -690
DOI:10.1007/s12190-020-01373-0
[本文引用: 1]
[20]
李诗琪 , 唐三一 . 具有群体防御的捕食-被捕食模型适应性进化分析
应用数学和力学 , 2023 , 44 3 ): 319 -332
Li S Q Tang S Y Adaptive evolution analysis of a predator-prey model with group defense
Applied Mathematics and Mechanics , 2023 , 44 3 ): 319 -332
[21]
Yao Y Liu L Dynamics of a predator-prey system with foraging facilitation and group defense
Communications in Nonlinear Science and Numerical Simulation , 2024 , 138
[本文引用: 1]
[22]
Turing A M The chemical basis of morphogenesis. Philosophical Transactions of the Royal Society of London
SerieB, Biological Sciences , 1952 , 237 641 ): 37 -72
[本文引用: 1]
[23]
Ali I Saleem M T Spatiotemporal dynamics of reaction-diffusion system and its application to turing pattern formation in a gray-scott model
Mathematics , 2023 , 11 6 ): Art 1459
[本文引用: 1]
[24]
Sgura I Bozzini B Lacitignola D Numerical approximation of oscillating Turing patterns in a reaction-diffusion model for electrochemical material growth
In AIP Conference Proceedings American Institute of Physics , 2012 , 1493 1 ): 896 -903
[25]
Kondo S Miura T Reaction-diffusion model as a framework for understanding biological pattern formation
Science , 2010 , 329 5999 ): 1616 -1620
DOI:10.1126/science.1179047
PMID:20929839
[本文引用: 1]
The Turing, or reaction-diffusion (RD), model is one of the best-known theoretical models used to explain self-regulated pattern formation in the developing animal embryo. Although its real-world relevance was long debated, a number of compelling examples have gradually alleviated much of the skepticism surrounding the model. The RD model can generate a wide variety of spatial patterns, and mathematical studies have revealed the kinds of interactions required for each, giving this model the potential for application as an experimental working hypothesis in a wide variety of morphological phenomena. In this review, we describe the essence of this theory for experimental biologists unfamiliar with the model, using examples from experimental studies in which the RD model is effectively incorporated.
[26]
Holmes E E Lewis M A Banks J E et al . Partial differential equations in ecology: Spatial interactions and population dynamics
Ecology , 1994 , 75 1 ): 17 -29
DOI:10.2307/1939378
URL
[本文引用: 1]
Most of the fundamental elements of ecology, ranging from individual behavior to species abundance, diversity, and population dynamics, exhibit spatial variation. Partial differential equation models provide a means of melding organism movement with population processes and have been used extensively to elucidate the effects of spatial variation on populations. While there has been an explosion of theoretical advances in partial differential equation models in the past two decades, this work has been generally neglected in mathematical ecology textbooks. Our goal in this paper is to make this literature accessible to experimentally ecologists. Partial differential equations are used to model a variety of ecological phenomena; here we discuss dispersal, ecological invasions, critical patch size, dispersal—mediated coexistence, and diffusion—driven spatial patterning. These model emphasize that simple organism movement can produce striking large—scale patterns in homogeneous environments, and that in heterogeneous environments, movement of multiple species can change the outcome of competition or predation.
[27]
Zhou Y Yan X P Zhang C H Turing patterns induced by self-diffusion in a predator-prey model with schooling behavior in predator and prey
Nonlinear Dynamics , 2021 , 105 4 ): 3731 -3747
DOI:10.1007/s11071-021-06743-2
[本文引用: 1]
[28]
肖江龙 , 宋永利 , 夏永辉 . 一个扩散模型中恐惧效应与集群行为协同诱导的时空动力学研究
数学物理学报 , 2024 , 44 6 ): 1577 -1594
[本文引用: 1]
Xiao J L Song Y L Xia Y H A study on the spatiotemporal dynamics induced by fear effects and clustering behavior in a diffusion model
Acta Math Sci , 2024 , 44 6 ): 1577 -1594
[本文引用: 1]
[29]
Kerner E H A statistical mechanics of interacting biological species
The Bulletin of Mathematical Biophysics , 1957 , 19 2 ): 121 -146
DOI:10.1007/BF02477883
URL
[本文引用: 1]
[30]
Jalilian Y Farshid M Hopf bifurcation and Turing instability in a cross-diffusion prey-predator system with group defense behavior
Computational Methods for Differential Equations , 2024 , 13 1 ): 294 -306
[本文引用: 1]
[31]
Tulumello E Lombardo M C Sammartino M Cross-diffusion driven instability in a predator-prey system with cross-diffusion
Acta Applicandae Mathematicae , 2014 , 132 1 ): 621 -633
DOI:10.1007/s10440-014-9935-7
URL
[本文引用: 1]
[32]
Wang H Liu P Pattern dynamics of a predator-prey system with cross-diffusion, Allee effect and generalized Holling IV functional response
Chaos Solitons Fractals , 2023 , 171
[本文引用: 1]
[33]
Han R Dey S Huang J et al . Spatio-temporal steady-state analysis in a prey-predator model with saturated hunting cooperation and chemotaxis
Acta Applicandae Mathematicae , 2024 , 191 1 ): Art 10
[本文引用: 1]
[34]
Cao J Ma L Hao P Bifurcation analysis in a modified Leslie-Gower predator-prey model with Beddington-DeAngelis functional response
Journal of Applied Analysis Computation , 2023 , 132 5 ): 3026 -3053
[本文引用: 2]
[35]
范示示 , 李海侠 , 路银豆 . 具有捕获项的 Beddington-DeAnglis 型捕食-食饵扩散模型的动力学分析
数学物理学报 , 2023 , 43 6 ): 1929 -1942
[本文引用: 1]
Fan S S Li H X Lu Y D The dynamical analysis of a diffusive predator-prey model with Beddington-DeAngelis type functional response incorporating prey capture
Acta Math Sci , 2023 , 43 6 ): 1929 -1942
[本文引用: 1]
[36]
李波 , 梁子维 . 一类具有 Beddington-DeAngelis 响应函数的阶段结构捕食模型的稳定性
数学物理学报 , 2022 , 42 6 ): 1826 -1835
[本文引用: 1]
Li B Liang Z W Dynamical stability in a stage-structured predator-prey system incorporating Beddington-DeAngelis type functional response
Acta Math Sci , 2022 , 42 6 ): 1826 -1835
[本文引用: 1]
Cross-diffusion mediated Spatiotemporal patterns in a predator-prey system with hunting cooperation and fear effect
1
2024
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
Variations and fluctuations of the number of individuals in animal species living together
1
1928
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
The orgins and evolution of predator-prey theory
1
1992
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
Further studies of interaction between predators and prey
0
1936
On competition between different species of graminivorous insects
0
1945
An analysis of models describing predator-prey interaction
0
1970
On the Volterra and other nonlinear models of interacting populations
1
1971
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
A predator-prey model with a Holling type I functional response including a predator mutual interference
1
2011
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
Dynamics of a predator-prey model with generalized Holling type functional response and mutual interference
1
2020
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
Global dynamics of a predator-prey system with density-dependent mortality and ratio-dependent functional response
1
2021
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
On the dynamics of predator-prey models with the Beddington-DeAngelis functional response
1
2001
... 捕食者与猎物之间的动态相互作用是生态学中促进种群进化和保护生物多样性的核心问题[1 ] . 自 1928 年 Vito Volterra 提出著名的捕食者-猎物系统[2 ] , 该系统在生物学和应用数学领域迅速被接纳[3 7 ] . 但是原始系统过于理想化, 未能充分体现出捕食者- 猎物间复杂的相互作用. 因此, 学者们提出了多种功能响应函数来改进对捕食者-猎物复杂关系的刻画. Seo 等[8 ] 采用 Holling I 型功能响应函数模拟捕食者干扰. Antwi-Fordjour 等[9 ] 建立了广义 Holling 型系统, 给出了 Hopf 分岔条件和猎物灭绝判据. Jiang 等[10 ] 揭示了比率依赖型系统可存在 Bogdanov-Takens 分岔等复杂动力学特征. Cantrell 等[11 ] 基于 Beddington-DeAngelis 功能响应函数推导了捕食者永久性存在和灭绝的条件. 其中 Beddington-DeAngelis 功能响应函数同时考虑了猎物密度和捕食者密度对捕食效率的影响, 并且具有更为合理的饱和项和捕食干扰项. 因此本文采用 Beddington-DeAngelis 功能响应函数来刻画捕食关系. 上述研究普遍考虑了较为复杂的功能响应函数, 其根本原理都是反映捕食者直接捕食猎物的机制. ...
Role of fear in predator-prey system with intraspecific competition
1
2020
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
具有恐惧效应的时滞捕食者-食饵模型
1
2023
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
A predator-prey model with fear effect and time delay
1
2023
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
Role of fear in a predator-prey model with Beddington-DeAngelis functional response
0
2019
Impact of fear effect on the growth of prey in a predator-prey interaction model
1
2020
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
Competition and coexistence in plant communities: Intraspecific competition is stronger than interspecific competition
1
2018
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
The mechanistic basis for higher-order interactions and non-additivity in competitive communities
0
2019
具有集群效应和种内竞争的双时滞捕食模型的稳定性与 Hopf 分支
1
2022
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
Stability and hopf bifurcation of a dual-delay predator-prey model with grouping effect and intra-specific competition
1
2022
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
The effect of the defensive strategy taken by the prey on predator-prey interaction
1
2020
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
具有群体防御的捕食-被捕食模型适应性进化分析
0
2023
Adaptive evolution analysis of a predator-prey model with group defense
0
2023
Dynamics of a predator-prey system with foraging facilitation and group defense
1
2024
... 事实上, 捕食者的存在也可能改变猎物的行为模式和生理, 其影响比直接捕食更深远[12 ] . 因此, 间接相互作用联合功能响应函数的相关模型形式引起了广泛关注. Wang 等[13 15 ] 将恐惧效应引入到捕食者-猎物系统中, 分析了恐惧效应对 Hopf 分岔的影响, 证明了恐惧效应与猎物出生率存在负相关关系. Adler 等[16 18 ] 研究了种内竞争对种群动态的影响, 其实验数据证明了种内竞争推动了生物多样性的发展, 对种群生态的发展具有深远影响. Souna 等[19 21 ] 考虑了猎物的群体防御行为, 揭示了群体防御对系统的稳定性和 Hopf 分岔有重要影响. 值得注意的是, 这些研究仅基于常微分方程, 考虑单一相互作用下的种群动力学. 因此本文引入空间维度建立反应扩散系统, 并综合考虑恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用. ...
The chemical basis of morphogenesis. Philosophical Transactions of the Royal Society of London
1
1952
... Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25 ] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用. ...
Spatiotemporal dynamics of reaction-diffusion system and its application to turing pattern formation in a gray-scott model
1
2023
... Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25 ] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用. ...
Numerical approximation of oscillating Turing patterns in a reaction-diffusion model for electrochemical material growth
0
2012
Reaction-diffusion model as a framework for understanding biological pattern formation
1
2010
... Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25 ] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用. ...
Partial differential equations in ecology: Spatial interactions and population dynamics
1
1994
... Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25 ] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用. ...
Turing patterns induced by self-diffusion in a predator-prey model with schooling behavior in predator and prey
1
2021
... Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25 ] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用. ...
一个扩散模型中恐惧效应与集群行为协同诱导的时空动力学研究
1
2024
... Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25 ] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用. ...
A study on the spatiotemporal dynamics induced by fear effects and clustering behavior in a diffusion model
1
2024
... Turing[22 ] 在 1952 年首次使用反应-扩散方程解释了生物表面图案的形成过程. 他发现在反应扩散系统中, 存在于扩散率差异较大的两种不同物质之间的反应和扩散过程可以通过自发破坏对称性而形成斑图. 此后斑图动力学在多个领域被广泛研究[23 25 ] . 生态系统中也存在扩散现象, 物种在空间上运动行为的复杂多样性, 会形成不同的种群分布[26 ] . Zhou 等[27 ] 研究了捕食者-猎物中自扩散诱导的图灵斑图, 该模型表现出的复杂的动力学行为有助于理解两个物种间的相互作用. Xiao 等[28 ] 研究了含自扩散和恐惧以及集群效应的捕食者-猎物模型的时空动力学. 但这些研究并没有考虑交叉扩散的作用. ...
A statistical mechanics of interacting biological species
1
1957
... 在前人研究自扩散影响的基础上, Kerner[29 ] 首次提出了交叉扩散. 与考虑扩散在物种内发生的自扩散系统不同, 含交叉扩散的系统能够描述捕食者与猎物物种之间的扩散. Jalilian 等[30 ] 研究了具有群体防御行为的交叉扩散捕食者-猎物系统中的 Hopf 分岔和 Turing 不稳定性, 检查了分岔周期解的稳定性和 Hopf 分岔的方向. Tulumello 等[31 ] 研究了含交叉扩散的捕食者-猎物反应扩散系统的图灵斑图的形成过程, 通过非线性分析和数值模拟, 预测且验证了斑图模式以及波的形状和速度. Wang 等[32 ] 研究了具有交叉扩散、Allee 效应和 Holling IV 型功能响应函数的捕食者-猎物系统的斑图动力学, 给出了系统 Turing 不稳定、Hopf 不稳定和 Turing-Hopf 不稳定的理论分析. Han 等[33 ] 提出了一种具有捕食者趋向性的反应扩散捕食者-猎物系统, 揭示了斑图的形成是由捕食者趋向性和捕食者扩散的共同作用驱动的. 目前, 对反应扩散系统的研究多局限于单一相互作用和功能响应函数的情形. 因此, 本文基于改进型 Leslie-Gower 系统, 考虑 Beddington-DeAngelis 功能响应函数, 研究恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用机制影响下系统的时空动力学, 力图深入探明实际捕食者-猎物系统的动力学行为. ...
Hopf bifurcation and Turing instability in a cross-diffusion prey-predator system with group defense behavior
1
2024
... 在前人研究自扩散影响的基础上, Kerner[29 ] 首次提出了交叉扩散. 与考虑扩散在物种内发生的自扩散系统不同, 含交叉扩散的系统能够描述捕食者与猎物物种之间的扩散. Jalilian 等[30 ] 研究了具有群体防御行为的交叉扩散捕食者-猎物系统中的 Hopf 分岔和 Turing 不稳定性, 检查了分岔周期解的稳定性和 Hopf 分岔的方向. Tulumello 等[31 ] 研究了含交叉扩散的捕食者-猎物反应扩散系统的图灵斑图的形成过程, 通过非线性分析和数值模拟, 预测且验证了斑图模式以及波的形状和速度. Wang 等[32 ] 研究了具有交叉扩散、Allee 效应和 Holling IV 型功能响应函数的捕食者-猎物系统的斑图动力学, 给出了系统 Turing 不稳定、Hopf 不稳定和 Turing-Hopf 不稳定的理论分析. Han 等[33 ] 提出了一种具有捕食者趋向性的反应扩散捕食者-猎物系统, 揭示了斑图的形成是由捕食者趋向性和捕食者扩散的共同作用驱动的. 目前, 对反应扩散系统的研究多局限于单一相互作用和功能响应函数的情形. 因此, 本文基于改进型 Leslie-Gower 系统, 考虑 Beddington-DeAngelis 功能响应函数, 研究恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用机制影响下系统的时空动力学, 力图深入探明实际捕食者-猎物系统的动力学行为. ...
Cross-diffusion driven instability in a predator-prey system with cross-diffusion
1
2014
... 在前人研究自扩散影响的基础上, Kerner[29 ] 首次提出了交叉扩散. 与考虑扩散在物种内发生的自扩散系统不同, 含交叉扩散的系统能够描述捕食者与猎物物种之间的扩散. Jalilian 等[30 ] 研究了具有群体防御行为的交叉扩散捕食者-猎物系统中的 Hopf 分岔和 Turing 不稳定性, 检查了分岔周期解的稳定性和 Hopf 分岔的方向. Tulumello 等[31 ] 研究了含交叉扩散的捕食者-猎物反应扩散系统的图灵斑图的形成过程, 通过非线性分析和数值模拟, 预测且验证了斑图模式以及波的形状和速度. Wang 等[32 ] 研究了具有交叉扩散、Allee 效应和 Holling IV 型功能响应函数的捕食者-猎物系统的斑图动力学, 给出了系统 Turing 不稳定、Hopf 不稳定和 Turing-Hopf 不稳定的理论分析. Han 等[33 ] 提出了一种具有捕食者趋向性的反应扩散捕食者-猎物系统, 揭示了斑图的形成是由捕食者趋向性和捕食者扩散的共同作用驱动的. 目前, 对反应扩散系统的研究多局限于单一相互作用和功能响应函数的情形. 因此, 本文基于改进型 Leslie-Gower 系统, 考虑 Beddington-DeAngelis 功能响应函数, 研究恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用机制影响下系统的时空动力学, 力图深入探明实际捕食者-猎物系统的动力学行为. ...
Pattern dynamics of a predator-prey system with cross-diffusion, Allee effect and generalized Holling IV functional response
1
2023
... 在前人研究自扩散影响的基础上, Kerner[29 ] 首次提出了交叉扩散. 与考虑扩散在物种内发生的自扩散系统不同, 含交叉扩散的系统能够描述捕食者与猎物物种之间的扩散. Jalilian 等[30 ] 研究了具有群体防御行为的交叉扩散捕食者-猎物系统中的 Hopf 分岔和 Turing 不稳定性, 检查了分岔周期解的稳定性和 Hopf 分岔的方向. Tulumello 等[31 ] 研究了含交叉扩散的捕食者-猎物反应扩散系统的图灵斑图的形成过程, 通过非线性分析和数值模拟, 预测且验证了斑图模式以及波的形状和速度. Wang 等[32 ] 研究了具有交叉扩散、Allee 效应和 Holling IV 型功能响应函数的捕食者-猎物系统的斑图动力学, 给出了系统 Turing 不稳定、Hopf 不稳定和 Turing-Hopf 不稳定的理论分析. Han 等[33 ] 提出了一种具有捕食者趋向性的反应扩散捕食者-猎物系统, 揭示了斑图的形成是由捕食者趋向性和捕食者扩散的共同作用驱动的. 目前, 对反应扩散系统的研究多局限于单一相互作用和功能响应函数的情形. 因此, 本文基于改进型 Leslie-Gower 系统, 考虑 Beddington-DeAngelis 功能响应函数, 研究恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用机制影响下系统的时空动力学, 力图深入探明实际捕食者-猎物系统的动力学行为. ...
Spatio-temporal steady-state analysis in a prey-predator model with saturated hunting cooperation and chemotaxis
1
2024
... 在前人研究自扩散影响的基础上, Kerner[29 ] 首次提出了交叉扩散. 与考虑扩散在物种内发生的自扩散系统不同, 含交叉扩散的系统能够描述捕食者与猎物物种之间的扩散. Jalilian 等[30 ] 研究了具有群体防御行为的交叉扩散捕食者-猎物系统中的 Hopf 分岔和 Turing 不稳定性, 检查了分岔周期解的稳定性和 Hopf 分岔的方向. Tulumello 等[31 ] 研究了含交叉扩散的捕食者-猎物反应扩散系统的图灵斑图的形成过程, 通过非线性分析和数值模拟, 预测且验证了斑图模式以及波的形状和速度. Wang 等[32 ] 研究了具有交叉扩散、Allee 效应和 Holling IV 型功能响应函数的捕食者-猎物系统的斑图动力学, 给出了系统 Turing 不稳定、Hopf 不稳定和 Turing-Hopf 不稳定的理论分析. Han 等[33 ] 提出了一种具有捕食者趋向性的反应扩散捕食者-猎物系统, 揭示了斑图的形成是由捕食者趋向性和捕食者扩散的共同作用驱动的. 目前, 对反应扩散系统的研究多局限于单一相互作用和功能响应函数的情形. 因此, 本文基于改进型 Leslie-Gower 系统, 考虑 Beddington-DeAngelis 功能响应函数, 研究恐惧效应、种内竞争、饱和效应、捕食干扰多种相互作用机制影响下系统的时空动力学, 力图深入探明实际捕食者-猎物系统的动力学行为. ...
Bifurcation analysis in a modified Leslie-Gower predator-prey model with Beddington-DeAngelis functional response
2
2023
... 根据文献 [34 ] 中的研究, 综合考虑改进的 Leslie-Gower 项和 Beddington-DeAngelis 功能响应函数, 能有效描述捕食者在不同种群密度下的捕食行为. 但并没有考虑捕食者-猎物之间广泛存在的相互作用和物种迁徙模式. 为了提高系统的普遍性和真实性, 在文献 [34 ] 的基础上, 加入恐惧效应、种内竞争等相互作用, 并进一步引入交叉扩散. 改进的系统模型如下 ...
... ] 中的研究, 综合考虑改进的 Leslie-Gower 项和 Beddington-DeAngelis 功能响应函数, 能有效描述捕食者在不同种群密度下的捕食行为. 但并没有考虑捕食者-猎物之间广泛存在的相互作用和物种迁徙模式. 为了提高系统的普遍性和真实性, 在文献 [34 ] 的基础上, 加入恐惧效应、种内竞争等相互作用, 并进一步引入交叉扩散. 改进的系统模型如下 ...
具有捕获项的 Beddington-DeAnglis 型捕食-食饵扩散模型的动力学分析
1
2023
... $\textbf{注 5.3}\quad$ 图4 到图6 斑图模式转变的解释: 根据文献 [35 ,36 ] 的研究, 捕食者对猎物的饱和效应本质上反映了捕食过程中的时间成本约束. 捕食者从捕获猎物到完成进食、消化都需消耗一定时间资源, 这导致其单位时间内可处理的猎物数量存在上限. 当 $b$ $b$
The dynamical analysis of a diffusive predator-prey model with Beddington-DeAngelis type functional response incorporating prey capture
1
2023
... $\textbf{注 5.3}\quad$ 图4 到图6 斑图模式转变的解释: 根据文献 [35 ,36 ] 的研究, 捕食者对猎物的饱和效应本质上反映了捕食过程中的时间成本约束. 捕食者从捕获猎物到完成进食、消化都需消耗一定时间资源, 这导致其单位时间内可处理的猎物数量存在上限. 当 $b$ $b$
一类具有 Beddington-DeAngelis 响应函数的阶段结构捕食模型的稳定性
1
2022
... $\textbf{注 5.3}\quad$ 图4 到图6 斑图模式转变的解释: 根据文献 [35 ,36 ] 的研究, 捕食者对猎物的饱和效应本质上反映了捕食过程中的时间成本约束. 捕食者从捕获猎物到完成进食、消化都需消耗一定时间资源, 这导致其单位时间内可处理的猎物数量存在上限. 当 $b$ $b$
Dynamical stability in a stage-structured predator-prey system incorporating Beddington-DeAngelis type functional response
1
2022
... $\textbf{注 5.3}\quad$ 图4 到图6 斑图模式转变的解释: 根据文献 [35 ,36 ] 的研究, 捕食者对猎物的饱和效应本质上反映了捕食过程中的时间成本约束. 捕食者从捕获猎物到完成进食、消化都需消耗一定时间资源, 这导致其单位时间内可处理的猎物数量存在上限. 当 $b$ $b$





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}